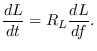Next: Larvae cycle Up: VECTRI manual Previous: Calibration
The malaria model presented is a grid cell distributed dynamical model and is referred to as VECTRI; the vector borne disease community model of the International Centre for Theoretical Physics, Trieste1. In as far as possible, the model physics and associated parameters are taken from the literature for the Anopheles gambiae complex and the Plasmodium falciparum malaria parasite. In the present version each location (grid cell) is independent, but the structure of the model will allow communication between grid cells such as vector flight or human population migration to be easily incorporated. The following sections describe the basic structure of the model, with emphasis placed on its novel aspects.
Since one goal of the model is its successful application regionally to
forecast epidemic outbreaks in malaria marginal zones in addition to
representing malaria transmission in endemic regions, it is
important to represent the delay between the rainy season onset and
the malaria season. Thus the model explicitly resolves the growth
stages of the egg-larvae-pupa cycle in addition to the gonotrophic and
the sporogonic cycles using an array of bins for each process, similar
to the LMM. The structure of the model is depicted schematically in
Fig.1. It shows the division of the larvae
life cycle (![]() ) into a number (
) into a number (![]() ) of discrete fractional bins
(
) of discrete fractional bins
(![]() ). The real number stored in each bin,
). The real number stored in each bin, ![]() , gives the larvae
density (per square metre) at a particular fractional growth stage
, gives the larvae
density (per square metre) at a particular fractional growth stage ![]() (where
(where ![]() ranges from 0 to 1). Oviposition results in eggs added
to the first bin and each timestep of the model larvae advance a
number of bins at a fractional growth rate
ranges from 0 to 1). Oviposition results in eggs added
to the first bin and each timestep of the model larvae advance a
number of bins at a fractional growth rate ![]() (units
(units
![]() ), depending on the local water temperature, until they reach
the final bin (representing
), depending on the local water temperature, until they reach
the final bin (representing ![]() ) and develop into adult mosquitoes.
The equations solved are thus the classic advection equation:
) and develop into adult mosquitoes.
The equations solved are thus the classic advection equation:
The vector status is also bin resolved, consisting of two properties:
the gonotrophic and sporogonic cycles. It is thus
represented as a two dimensional array
![]() . All
vectors in the first gonotrophic bin
. All
vectors in the first gonotrophic bin
![]() are in
meal-searching mode, and once a meal is obtained, the vectors advance in terms of
the egg development state at a rate
are in
meal-searching mode, and once a meal is obtained, the vectors advance in terms of
the egg development state at a rate
![]() related to ambient
temperature until the final bin is reached (using the advection
equation similar to eqn.
related to ambient
temperature until the final bin is reached (using the advection
equation similar to eqn. ![[*]](crossref.png) ). At this point the vector
lays a new raft of eggs and is recycled to the first meal-searching
bin.
). At this point the vector
lays a new raft of eggs and is recycled to the first meal-searching
bin.
Each timestep, parasite transmission may occur to a proportion of the
biting vectors, and the status of these vectors will subsequently
additionally progress in the sporogonic dimension, with the rate
![]() again determined by temperature. Once vectors reach the
final bin they are infective to humans and remain so until death. A
third array of bins maps the status of the disease in the human host
(
again determined by temperature. Once vectors reach the
final bin they are infective to humans and remain so until death. A
third array of bins maps the status of the disease in the human host
(![]() ) population (dimension
) population (dimension ![]() ), with the first bin representing
the uninfected population. The model does not include age or immunity
factors, which is the subject of present model development.
), with the first bin representing
the uninfected population. The model does not include age or immunity
factors, which is the subject of present model development.
Thus, while the model introduces new relationships regarding the surface hydrology and the explicit interaction between vector and host, the underlying numerical structure is similar to that employed in the LMM [#!hoshen:04!#]. A timestep of one day is used to integrate the model equations, although a shorter timestep could be used if input data (temperature/rainfall) are available on these timescales, and the advection equation is solved using a simple upstream numerical scheme.
The model can be flexibly integrated using a wide range of horizontal resolutions. Since the model presently does not permit vectors to move to neighbouring grid cells, the model resolution is limited to an upper (finest) level on the order of 1-5 km; an indicated range below which mosquito movement can become significant [#!gillies:61!#,#!rowley:68!#,#!thomson:95!#]. Integrating the model at O(10km) resolution is desirable, even if corresponding observational data of clinically proven malaria cases is available only on coarser scale health districts. This is because malaria transmission is a highly nonlinear function of its drivers such as climate and land surface and thus it is preferable to use highest possible spatial resolution inputs of population density, land surface and weather to account for this, while the model output is subsequently aggregated to the spatial scale at which health data is available for comparison.
The following section describes the details of the parametrizations used to describe the temperature-sensitive progression rates and details the new developments in the VECTRI model concerning the representation of the human population density, which impacts the biting rates, and the treatment of surface hydrology which determines the overall vector number. To enable VECTRI to be used in a multi-model ensemble approach to assess model uncertainties, multiple parametrization choices are incorporated to facilitate this. All the constants are listed in table 1.