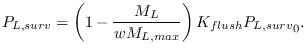Next: Gonotrophic cycle Up: VECTRI Description Previous: Larvae cycle
The mortality rate of larvae is an important factor for transmission,
and is strongly temperature dependent [#!kirby:09!#].
The VECTRI model sets a base daily survival rate for larvae
![]() taken from [#!ermert:11a!#], slightly lower
than the values given by [#!hoshen:04!#,#!bomblies:08!#]. Development of
larvae is negatively affected by larvae over-population due
to competition for resources [#!armstrong:61!#]. This is
incorporated in VECTRI by reducing survival rate proportionally by a
factor related to resource constraints:
taken from [#!ermert:11a!#], slightly lower
than the values given by [#!hoshen:04!#,#!bomblies:08!#]. Development of
larvae is negatively affected by larvae over-population due
to competition for resources [#!armstrong:61!#]. This is
incorporated in VECTRI by reducing survival rate proportionally by a
factor related to resource constraints:
Flushing of larvae by heavy rainfall has been suggested to be an
important cause of larva mortality [#!martens:95b!#,#!thomson:05!#].
Using an artificial pond apparatus [#!paaijmans:07a!#], a 17.5%
daily mortality rate of first stage larvae reducing to 4.8% for forth
stage larvae was estimated, although the authors state that
these figures could represent an underestimate
due to the symmetry of the pond apparatus and the lack of sampling of
more extreme rainfall amounts during the experiment campaign. On the
other hand, vegetation in natural pools and the apparent ability for
Anopheles gambiae larvae to take avoidance measures to avoid
flushing could imply a lower flushing rate [#!muirhead:58!#]. In
order take the simplest possible relationship in VECTRI, flushing rate
is represented as exponential function of rate rate, and is related
linearly to the larvae fractional growth state ![]() :
:
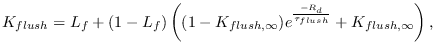 |
(4) |
| parameter | definition | default | units |
| nlarv_scheme | larvae water temperature scheme (1,2,3) | 1 | |
| 1=ermert:2011a,2=jepsen47,3=bayoh lindsey 2003 | |||
| nsurvival_scheme | mosquito survival temperature scheme (1,2,3) | 2 | |
| 1=MartinsI,2=MartinsII,3=Bayoh(not yet!) |