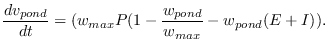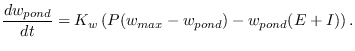Next: internal pond calculation, revised Up: w_pond Previous: external pond calculation
The net aggregated fractional coverage by temporary pools ![]() is derived from a simple water balance model. Ponds are replenished
by surface runoff
is derived from a simple water balance model. Ponds are replenished
by surface runoff ![]() , while infiltration
, while infiltration ![]() (seepage) and
evaporation
(seepage) and
evaporation ![]() and pond overflow reduce their water content.
An important parameter is the maximum coverage of the temporary ponds
and pond overflow reduce their water content.
An important parameter is the maximum coverage of the temporary ponds
![]() , which described the extent of depressions that could
potentially become water filled at the peak of a wet season, which are
often small in scale, with total catchments of the gully systems
studies in Niger in the HAPEX-SAHEL experiment measuring 0.2 km
, which described the extent of depressions that could
potentially become water filled at the peak of a wet season, which are
often small in scale, with total catchments of the gully systems
studies in Niger in the HAPEX-SAHEL experiment measuring 0.2 km![]() .
Presently is assumed that the runoff
.
Presently is assumed that the runoff ![]() that fills the ponds mostly
falls within these depressions and thus the run off is set to
that fills the ponds mostly
falls within these depressions and thus the run off is set to
![]() . Thus sub-surface infiltration occurring within the
depressions is also considered a water source for temporary pools
[#!desconnets:97!#]. Future developments will introduce improved runoff
treatment accounting for soil texture and slope.
. Thus sub-surface infiltration occurring within the
depressions is also considered a water source for temporary pools
[#!desconnets:97!#]. Future developments will introduce improved runoff
treatment accounting for soil texture and slope.
The source from precipitation is balanced by evaporative, infiltration
and overflow losses. In high resolution simulations of surface
hydrology in Niger, [#!gianotti:09!#] found overflow losses to be
approximately 20% of total losses, more than three times the losses
due to evaporation. It should also be noted that overflow losses in
field campaigns are difficult to measure and thus are often
incorporated in the infiltration, which is calculated as a residual in
the water balance calculation. Losses through pond overflow are
assumed to increase linearly with pond fraction in VECTRI, achieved by
scaling the runoff by a factor
![]() . Once the
pond fraction reaches its maximum, all surface runoff overflows and is
lost. Infiltration losses
vary substantially depending on soil texture and life-scale of the
pond in question. Often the infiltration is a highly nonlinear
function of water body extent, since silting may significantly reduce
infiltration in the lowest part of longer-lived or semi-permanent
pools [#!desconnets:97!#]. This results in a fast initial decay
after rain events due to high infiltration rates at the pool edges,
followed by a slower decay, while temporary, shorter lived water
bodies tend to have more uniform infiltration rates. These rates can
be very high, exceeding 600 mm day
. Once the
pond fraction reaches its maximum, all surface runoff overflows and is
lost. Infiltration losses
vary substantially depending on soil texture and life-scale of the
pond in question. Often the infiltration is a highly nonlinear
function of water body extent, since silting may significantly reduce
infiltration in the lowest part of longer-lived or semi-permanent
pools [#!desconnets:97!#]. This results in a fast initial decay
after rain events due to high infiltration rates at the pool edges,
followed by a slower decay, while temporary, shorter lived water
bodies tend to have more uniform infiltration rates. These rates can
be very high, exceeding 600 mm day![]() [#!martin:03!#]. Presently
the VECTRI model simply sets a fixed constant infiltration rate per
unit pond area.
[#!martin:03!#]. Presently
the VECTRI model simply sets a fixed constant infiltration rate per
unit pond area.
Combining these factors, the volume of water ![]() in ponds
per unit area thus evolves as
in ponds
per unit area thus evolves as
![[*]](crossref.png) is
integrated using a fully implicit solution. An example evolution of
the fractional pond coverage for a site near Bobo Dioulasso is given
in Fig. 4, which shows that the simple empirical
approach mimics the pond evolution modelled by high resolution
hydrological models for sites in Niger [#!bomblies:08!#] and ponds
modelled in Senegal [#!soti:10!#]. The present empirical formulation
is similar to the approaches of [#!alonso:11!#,#!eckhoff:11!#]. It is
seen that at the fringes of the rainy season, puddles and small ponds
have limited longevity on the order of a few days, implying that they
are unsuitable for vector breeding. VECTRI represents
the bulk behaviour of ponds, rather than the ultra-high resolution
model of [#!gianotti:09!#] which individual models puddles at the 10
m scale and thus can model the lifetime of ponds as a function of
their explicit size (see their figure 7). Obviously, the linear
relation between rainfall and the growth of potential breeding sites
in parameter
is
integrated using a fully implicit solution. An example evolution of
the fractional pond coverage for a site near Bobo Dioulasso is given
in Fig. 4, which shows that the simple empirical
approach mimics the pond evolution modelled by high resolution
hydrological models for sites in Niger [#!bomblies:08!#] and ponds
modelled in Senegal [#!soti:10!#]. The present empirical formulation
is similar to the approaches of [#!alonso:11!#,#!eckhoff:11!#]. It is
seen that at the fringes of the rainy season, puddles and small ponds
have limited longevity on the order of a few days, implying that they
are unsuitable for vector breeding. VECTRI represents
the bulk behaviour of ponds, rather than the ultra-high resolution
model of [#!gianotti:09!#] which individual models puddles at the 10
m scale and thus can model the lifetime of ponds as a function of
their explicit size (see their figure 7). Obviously, the linear
relation between rainfall and the growth of potential breeding sites
in parameter
Presently, the two unknowns ![]() and
and ![]() in the framework are
set using a simple Monte Carlo suite of station data integrations in a
subset of locations to minimize
in the framework are
set using a simple Monte Carlo suite of station data integrations in a
subset of locations to minimize ![]() errors compared to field data.
An example sensitivity integration is shown in
Fig. 5, which is a integration conducted for Bobo
Dioulasso using station data to drive the model (see below for
experimental set up details). It is seen that transmission intensity
increases with
errors compared to field data.
An example sensitivity integration is shown in
Fig. 5, which is a integration conducted for Bobo
Dioulasso using station data to drive the model (see below for
experimental set up details). It is seen that transmission intensity
increases with ![]() and
and ![]() as expected, since these increase
the pond coverage for a given rainrate, while increasing the loss rate
acts in the opposite direction. For a given station there are a range
of reasonable parameter values, with the present parameter settings
chosen using a small number of locations in west Africa.
Nevertheless, the physically based framework facilitates future
improvement currently underway, which will include direct validation
of the revised hydrological model constants using in situ and remotely
sensed data.
as expected, since these increase
the pond coverage for a given rainrate, while increasing the loss rate
acts in the opposite direction. For a given station there are a range
of reasonable parameter values, with the present parameter settings
chosen using a small number of locations in west Africa.
Nevertheless, the physically based framework facilitates future
improvement currently underway, which will include direct validation
of the revised hydrological model constants using in situ and remotely
sensed data.
Finally, it is recalled that the pond dimension limits larvae
mortality rates through the availability of breeding sites governed in
eqn. . This is an approximation of the net affect
of crowding which leads to higher mortality rates, longer development
times and smaller adults [#!gimnig:02!#], which in turn have a
competative disadvantage [#!takken:98b!#]. The biomass is considered
to be distributed equally through all available breeding sites and
variability between breeding sites in neglected, supported by
[#!munga:06!#] who noted that females avoid ponds that are
overcrowded with existing larvae.
In addition to pond dimension the other important parameter of water
bodies is the temperature of the water near the surface. The Depinay
model [#!depinay:04!#] developed a complex empirical function for
water temperature as a function of ambient relative humidity and water
body size. As the VECTRI model is applied regionally, specific
information about individual water body size can not be included. The
temperature in shallow ponds and puddles is homogeneous to a good
approximation and is often one or two degrees warmer than the air
temperature [#!paaijmans:08b!#,#!paaijmans:08a!#]. VECTRI therefore
assumes that the temperature of pools ![]() to have a fixed offset
relative to the air temperature. The default value adopted is a
positive offset of 2 K, however, in hot locations it is likely that
vector will preferentially choose shaded breeding locations and a
lower or even negative offset may be more appropriate. If accurate
gridded weather information for wind and surface radiation were
available, this aspect of the model could be potential improved
implementing a single energy balance model along the lines of
[#!jacobs:08!#,#!paaijmans:08c!#]. While larger permanent water bodies
such as lakes and rivers can have complex stratification of the
vertical temperature profile, as discussed above, larvae development
occurs mostly in the shallow waters and pools that form on the
lake/river boundaries and thus the temperature relation for the
permanent water fraction is treated in the same way as the temporary
ponds.
to have a fixed offset
relative to the air temperature. The default value adopted is a
positive offset of 2 K, however, in hot locations it is likely that
vector will preferentially choose shaded breeding locations and a
lower or even negative offset may be more appropriate. If accurate
gridded weather information for wind and surface radiation were
available, this aspect of the model could be potential improved
implementing a single energy balance model along the lines of
[#!jacobs:08!#,#!paaijmans:08c!#]. While larger permanent water bodies
such as lakes and rivers can have complex stratification of the
vertical temperature profile, as discussed above, larvae development
occurs mostly in the shallow waters and pools that form on the
lake/river boundaries and thus the temperature relation for the
permanent water fraction is treated in the same way as the temporary
ponds.